Here we report the discovery of a new armoured dinosaur from the early Late Cretaceous of Argentina, recovered phylogenetically using various datasets either as a basal thyreophoran or a stem ankylosaur, closely related to Scelidosaurus.
It bears unusual anatomical features showing that several traits traditionally associated with the heavy Cretaceous thyreophorans did not occur universally.
Jakapil kaniukura gen.
Jakapil also shows that early thyreophorans had a much broader geographic distribution than previously thought.
It is a member of an ancient basal thyreophoran lineage that survived until the Late Cretaceous in South America.
Although most thyreophorans are grouped within Eurypoda (Ankylosauria + Stegosauria), basal thyreophorans show unresolved phylogenetic placements.
The most representative of these forms are Scutellosaurus, Emausaurus and Scelidosaurus, from the Lower Jurassic of the USA, Germany and England, respectively.
Both Scutellosaurus and Scelidosaurus represent part of the early locomotor diversity of early ornithischians, with obligate quadrupedality evolving in later and larger ankylosaurs and stegosaurs9.
We present here a new thyreophoran genus and species from the Late Cretaceous of South America.This new thyreophoran discovery from the LBPA provides new information about the structure of the early Late Cretaceous North Patagonian communities and the role of the still poorly known ornithischian component.
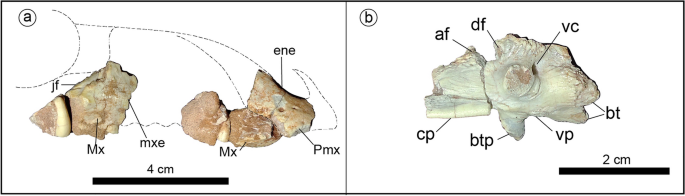
Holotype of Jakapil kaniukura (MPCA-PV-630), skull bones.(a) Skull bones in right lateral view (dashed contours based on Scelidosaurus10); (b) basisphenoid in left lateral view.
Holotype of Jakapil kaniukura (MPCA-PV-630), lower jaw bones.(a) left mandible in lateral view; (b) left mandible in lateral view, interpreted bone contours; (c) left mandible in medial view; (d) left mandible in medial view, interpreted bone contours; (e) right surangular in lateral view (mirrored); (f) transversal section of the posterior half of the left mandible, cranial view; (g) articular bone in occlusal view; (h) predentary bone in occlusal view.
Holotype of Jakapil kaniukura (MPCA-PV-630), teeth.Holotype of Jakapil kaniukura (MPCA-PV-630), postcranial bones.Speculative silhouette showing preserved elements (a); osteoderm distribution is speculative and partial to show non-osteodermal elements); dorsal vertebra elements in dorsal (b), right lateral (c) and anterior (d,e) views; sacral vertebra in left lateral view (f); mid-caudal vertebra in left lateral view (g); fragment of the mid-shaft of a dorsal rib in posterior view (the enlarged, broken posterior edge is highlighted (h); expanded distal ends of two dorsal ribs (i); left scapula in lateral view (j); right scapula in lateral view (k); right coracoid in lateral view (l); left and right humeri in anterior view (m); probable right ulna in lateral view (n); metacarpals, non-ungual and ungual phalanx in dorsal views (o); left femur elements in anterior view (p); proximal end of the right fibula in lateral view (q); distal end of the left tibia in anterior view (r); ischial elements in side view (s); cervical osteoderms in dorsal view (t), flat scutes in dorsal view (u), spine-like osteoderm in side view (v) and ossicle in dorsal view (w).
The genus, Jakapil (Ja-Kapïl: shield bearer), comes from the ‘gananah iahish’, Puelchean or northern Tehuelchean language.MPCA-PV-630 is a partial skeleton of a subadult individual (see Supplementary Information) that preserves fragments of some cranial bones (premaxilla, maxilla and basisphenoid), approximately 15 partial teeth and fragments, a nearly complete left lower jaw plus an isolated surangular, 12 partial vertebral elements, a complete dorsal rib and fifteen rib fragments, a partial coracoid, a nearly complete left scapula, a partial right scapula, two partial humeri, a possible partial right ulna, a complete and a partial metacarpal bone, three ischial and two femoral fragments, the distal end of a right tibia, the proximal end of a right fibula, three pedal phalanges, and more than forty osteoderms.Upper beds of the Candeleros Formation, early Late Cretaceous (Cenomanian, ~ 94–97 My, see16, and references therein), locality of Cerro Policía, Río Negro Province, North Patagonia, Argentina (Suppl. Fig. 1).Jakapil differs from all other thyreophorans in having: a large, ventral crest on the posterior half of the lower jaw, which is composed of the dentary, the angular and the splenial (medially hidden by the crest); a dorsomedially directed process in the short retroarticular process; leaf-shaped tooth crowns with a prominent mesial edge on their labial surface; maxillary and dentary tooth crowns differ from each other in their apical contour, the former being pointed and strongly asymmetrical, and the latter slightly curved distally with a more rounded and less asymmetrical contour; elongated (articular surface almost or completely beyond the posterior centrum face) and slender (width of less than a half postzygapophyses length) postzygapophyses in dorsal vertebrae; a strongly reduced humerus relative to the femur (proximal humeral width smaller than distal femoral width, see Supplementary Information), with a deep proximal fossa distally delimited by a curved ridge; a very large fibula relative to the femur (anteroposterior length of the proximal end almost comparable to the distal width of the femur); flattened and thin disk-like postcranial osteoderms.Jakapil is a small thyreophoran dinosaur (the subadult holotype is estimated to have been less than 1.5 m in body length and to have weighed 4.5–7 kg; see Supplementary Information, femoral description), with several novelties for a thyreophoran dinosaur.
The posterior half of the short lower jaw (Fig. 2a–f) is strongly dorsoventrally expanded, resembling the general shape of the heterodontosaurid33 and basal ceratopsian jaws34.
Anteriorly, the dentary oral margin is subhorizontal in lateral view (Fig. 2a–d, D), unlike the strongly downturned line of most thyreophorans30,37.
The glenoid fossa is roughly aligned with the tooth row in lateral view (Fig. 2a–d).
This process is absent in all other thyreophorans 9,10,35,36.
The tooth crowns are leaf-shaped as in basal ornithischian and thyreophorans10,28,29,38 (Fig. 3).The mesial edge of the labial surface in the maxillary and dentary tooth crowns is prominent as in Scelidosaurus10, and ends distally in a denticle-like structure in Jakapil (Fig. 3, me).
The diapophyses are laterodorsally directed almost 40° from the horizontal (Fig. 4d, di), at a lower angle than in stegosaurs28 and most ankylosaurs29, unlike the horizontal processes of basal ornithischians38.
An isolated mid-caudal vertebra shows an equidimensional centrum in lateral view, with concave, oval articular surfaces (Fig. 4g).
Some rib fragments with expanded (though broken) posterior edges suggest the presence of intercostal bones (Fig. 4h, ebr), as in Scelidosaurus39, Huayangosaurus43,44, some ankylosaurids45 (and references therein) and some basal ornithopods46.
The partial humeri (Fig. 3m) are strongly reduced in size, with overall limb proportions resembling those of basal ornithischians3,38 and several theropods47.
A possible proximal end of the ulna (Fig. 4n) resembles that of other basal ornithischians, though more strongly laterally compressed.
The preserved femoral pieces (Fig. 4p) resemble those of basal ornithischians38,39.
At least five osteoderm types are preserved in the holotype of Jakapil.The phylogenetic analysis using the matrix of Soto-Acuña et al.5 recovers Jakapil within Thyreophora, as the sister taxon of Ankylosauria (Fig. 5).The branch support for the basal thyreophorans is considerably lower than that obtained by Soto-Acuña et al.5, although the support of Stegosauria and some less inclusive eurypodan clades is slightly better (ceratopsians and pachycephalosaurs also show a lower support).
The Jakapil autapomorphies in this analysis are: ventrally orientated basipterygoid processes (char. 134; shared with Agilisaurus, Hypsilophodon, Zalmoxes, Tenontosaurus, Dryosaurus, Liaoceratops, Yamaceratops, Leptoceratops, Bagaceratops and Protoceratops); lateral orientation of the basipterygoid process articular facet (char. 136; shared with Homalocephale, Prenocephale, Stegoceras and Yinlong); a straight dentary tooth row in lateral view (char. 166; shared with the ornithischians Lesothosaurus, Eocursor, Scutellosaurus, Pinacosaurus, Euoplocephalus, heterodontosaurids and neornithischians); the presence of a ventral flange on the dentary (char. 170; shared with Psittacosaurus, Yamaceratops and Protoceratops); a well-developed coronoid process (char. 174; shared with heterodontosaurids and neornithischians); a surangular length of more than 50% the mandibular length (char. 183; shared with Stegoceras, Psittacosaurus, Yinlong, Chaoyangsaurus and Hualianceratops); less than 15 dentary teeth (char. 204; shared with heterodontosaurids, Gasparinisaura, Hypsilophodon, Wannanosaurus, Tenontosaurus, Dryosaurus and ceratopsians); apicobasally tall and blade-like cheek teeth crowns (char. 205; shared with Laquintasaura, Psittacosaurus, Yinlong, Chaoyangsaurus and Hualianceratops).
Alternative phylogenetic analyses using the data matrices of Maidment et al.4, Norman6 and Wiersma and Irmis8 recover Jakapil as the sister taxon of Eurypoda (Stegosauria + Ankylosauria) and as a basal ankylosaur, respectively (see Supplementary Information).
Being recovered either as an ankylosauromorph or a stem-eurypodan, Jakapil is closely related to Scelidosaurus in all analyses.
The discovery of Jakapil in the Cenomanian of Argentina shows a completely new thyreophoran lineage for the Southern Hemisphere.The new taxon shares many features with basal ornithischians and thyreophorans (even with ankylosaurids, see Supplementary Information), but also bears several novelties.
The relatively short mandible of Jakapil (Fig. 2a–d), with a large adductor fossa, extensively ornamented surangular, and a well-developed coronoid eminence (even higher than that of Scelidosaurus), resembles that of heterodontosaurids33 and basal ceratopsians34, suggesting a quite strong bite for a thyreophoran36,53.
The wear facets of Jakapil (Fig. 3, dwf, mwf) indicate dental occlusion; they are larger than those of most basal thyreophorans and stegosaurs, resembling those of the adult lectotype of Scelidosaurus10, ankylosaurs, ceratopsids and hadrosaurids36,54.
In Jakapil, the wear on functional cheek teeth is large and high angled, and seems largely distributed along the tooth rows, from the anteriormost teeth backward, as in heterodontosaurids and cerapodans33,54.
On the whole, it is probable that Jakapil had a masticatory system that was more efficient than the early thyreophorans in processing vegetation.
By contrast, the straight, narrow snout of Jakapil suggests a different feeding strategy, not cutting leaves but selecting elements that require harder processing36.
Thus, Jakapil expands the record of herbivorous vertebrates from the Kokorkom paleodesert, complementing this trophic level that contains the lepidosaur Priosphenodon20.
The predentary bone is the first known for a basal thyreophoran.Despite the variably complete lower jaws in Scutellosaurus9, Emausaurus42, Scelidosaurus10, “Tatisaurus”55 and “Bienosaurus”56, a predentary bone is absent.
The presence of an ossified predentary bone in Jakapil contrasts with that hypothesis.
Its plesiomorphic morphology mainly resembles that of the basal ornithischian Lesothosaurus31.
Several features of the predentary, such as the large ventral process, are shared with some basal ceratopsians34.
However, this process is usually very robust and proportionally larger that the lateral processes in basal ceratopsians when comparing with Jakapil.
In Jakapil, the glenoid fossa is not medially extended, and the pointed dorsomedial process arises from the retroarticular process.
However, the presence of a medial process in Jakapil may suggest a new placement of the pterygoid musculature, as in birds.
The armour of Jakapil is also peculiar.Almost all the recovered osteoderms are extremely low, unlike those of basal thyreophorans (Fig. 4r,s,u).
The Morphotype A osteoderms of Scutellosaurus62 are very low and bear a central keel, being roughly similar to the disk-shaped osteoderms of Jakapil (Fig. 4s; although the keel of the osteoderms in the latter is smoother).
Larger scutes in Jakapil show twice the radial extension of those of Scutellosaurus.
Some ankylosaurs, such as Gastonia52 and Gargoyleosaurus51, bear depressed plate-like osteoderms (with or without a low, sharp keel) resembling those of Jakapil.
In Scelidosaurus, the osteoderms develop a strong keel48, much higher than that seen in the Jakapil osteoderms.
By contrast, large, high-keeled osteoderms and spikes are rare in Jakapil.
Moreover, the mixture of plesiomorphic, stegosaurian and ankylosaurian characters of Jakapil may also suggest a basal phylogenetic placement (outside Eurypoda) for this taxon (contra6).
In addition, the incorporation of Jakapil into the data matrices of Soto-Acuña et al.5, Norman6, Maidment et al.4, and Wiersma and Irmis8 generates a general decrease in branch support.
This demonstrates that the early diversification of thyreophorans is still poorly understood due to their poor Lower-Middle Jurassic fossil record and the scarcity of Gondwanan material4,11, and may explain the ambiguous phylogenetic placement of Jakapil and the early thyreophorans2,4,6.
In Jakapil, the relative dimensions of the forelimb, hind limb, and cranial remains (Fig. 4a) bear a greater resemblance to those of the bipedal theropods47, basal ornithischians38 and heterodontosaurids33 than thyreophorans.
A comparison of the limb elements of some thyreophorans (Suppl. Fig. 4) shows the strong reduction in size of the humerus in Jakapil.Considering a reconstruction of the elements based on Scelidosaurus (the nearest taxon to Jakapil in all phylogenetic analyses), Scutellosaurus (a basal form) and Jinyunpelta (an ankylosaur), the reduction in size is evident.
Despite the incompleteness of the material, we quantified this reduction comparing the proximal humeral (PHW) and the distal femoral widths (DFW; the distal end of the femur in Jakapil was measured in the only well-preserved transversal section, although this is not the most distal).
The proximal humeral width/distal femoral width ratio (HFR) is lower in the basal taxa (Jakapil, Scutellosaurus and Scelidosaurus) with respect to the ankylosaurs Jinyunpelta and Euoplocephalus, showing a widening of the humerus in the quadrupedal taxa reaching a comparable width (ratio ~ 1).
Such widening in the proximal humeral end is evident in the lack of fit of the Jakapil bones in the Jinyunpelta proportions, also suggesting limb proportions more similar to those of basal forms.
Moreover, the incomplete distal end of the femur in Jakapil allows even smaller values of the HFR ratio (and of the humerus size).
Regardless of the unknown humeral length, the lack of a robust humerus in Jakapil allows us to reject a fully quadrupedal stance like that of the heavily built ankylosaurs.
In summary, the overall limb dimensions and estimations (with forelimb and olecranon process both reduced63), and the femoral anatomical similarities to the basal ornithischians and thyreophorans39 (e.g., large trochanters and a non-columnar element) suggest a bipedal stance in the specimen.However, the incompleteness of the remains demands caution to define the stance of Jakapil.
To make more complex the scenery, Jakapil still retains quadruped-associated features, such as a probable anterolateral process in the ulna, and stout metacarpals63 (and references therein).
The Early Jurassic thyreophoran record consists of basal forms known from several continents showing a Pangean distribution.Some recent papers recover the unarmoured ornithischians Lesothosaurus (Hettangian-Sinemurian of South Africa and Lesotho; see3,31) and Laquintasaura (Hettangian of Venezuela; see3,4,65) as Gondwanan basal members of Thyreophora.
The extensive distribution of armoured basal thyreophorans (excluding both Lesothosaurus and Laquintasaura) across the northern landmasses during the Early Jurassic shows a rapid diversification after the origin of the clade.However, basal thyreophoran remains from Gondwana known from the Middle Jurassic of Niger66, along with problematic material from the Lower Jurassic (Sinemurian-Pliensbachian) of India (see55,67 and references therein), suggest a more extensive distribution for the early thyreophorans.
The presence of the Middle Jurassic basal stegosaur Isaberrysaura from the Bajocian of Argentina68,4 and the stegosaurid Adratiklit from the Bathonian-Callovian of Morocco4 depict a distribution of the early stegosaurs that extends into southern landmasses.
The Middle Jurassic thyreophoran fossil record also includes the stegosaur Loricatosaurus from the Callovian of England and France, the ankylosaurs Sarcolestes and ‘Cryptosaurus’ from the Callovian of England, the ankylosaur Spicomellus from the Bathonian-Callovian of Morocco15, the ankylosaur ‘Tianchisaurus’ from the Callovian of China, and indeterminate remains from Europe and Asia (see4,67 and references therein).
On the whole, the Pangean distribution of early thyreophorans across the Early-Middle Jurassic makes it difficult to recognize a source area for Thyreophora and the most inclusive clades within it.
In this context, Jakapil not only increases the poor Gondwanan record of thyreophorans, but also establishes a theoretical framework for Gondwanan basal thyreophoran evolution and distribution (Fig. 5).Whether Jakapil is recovered as an ankylosaur or a non-eurypodan thyreophoran, it is closely related to Scelidosaurus (see Supplementary Information).
The presence of a basal thyreophoran in the early Late Cretaceous of South America shows that an ancient Gondwanan lineage of early thyreophorans evolved independently from those of the Northern Hemisphere, whose relationships have to be traced during the Early-Middle Jurassic Pangean rupture and the consequent isolation of Gondwana and later South America.
Accordingly, early Gondwanan thyreophorans survived a long time after the Pangean breakup, whereas the northern early thyreophorans seem to have gone extinct by the Middle Jurassic.
The persistence of a Gondwanan lineage of thyreophorans finds support in new thyreophoran remains recovered from the Lower Cretaceous rocks of the Bajada Colorada Formation (Berriasian-Hauterivian), also from the Argentinian Patagonia (Neuquén Province).
This clade includes Antarctopelta (Campanian-Maastrichthian of Antarctica), Stegouros (Campanian-Maastrichthian of South America), and the traditionally basal ankylosaur Kunbarrasaurus (Albian-Cenomanian of Australia)5.
Despite their extensive Gondwanan distribution during the Cretaceous, anatomical differences with Jakapil are remarkable.
Even more, ankylosaurian features present in Jakapil were recognized as convergent with ankylosaurids within Euankylosauria (e.g., a straight dentary tooth row in lateral view, a small diastema on the dentary, a shallow symphysis, scapular blade shape; see Supplementary Information) rather than with parankylosaurs. Therefore, a close phylogenetic relation between them is unlikely. Until more records contribute to the understanding of Gondwanan thyreophorans, both Jakapil and parankylosaurs belong to two different lineages. This shows that Gondwanan thyreophorans were a diverse clade with morphologically disparity..
On the other hand, further work will help to fill the extensive gap between the early thyreophorans and the Cretaceous remains from South America. Recent research is reflected in an increase in the thyreophoran fossil record from South America, with all its implications for thyreophoran evolution11,12,13,14,69,70; and this paper).The discovery of Jakapil not only supports the presence of a new Gondwanan lineage of early thyreophoran dinosaurs that persisted in Gondwana for a long time, but has also brought to light the importance of the Gondwanan fossil record in the study of the origin and evolution of dinosaurs (and other clades).
We used the dataset of Soto-Acuña et al.5, that comprises a broad sample of ornithischians suitable to test the phylogenetic position of Jakapil, a specimen with a complex mixture of features complete enough to include both several outgroups (non-thyreophoran ornithischians) and also both groups of thyreophorans.Basal ThyreophoraJ
Google Scholar .
Google Scholar .
Postcranial anatomy of Lesothosaurus diagnosticus (Dinosauria: Ornithischia) from the Lower Jurassic of southern Africa: Implications for basal ornithischian taxonomy and systematics.
Google Scholar .
Google Scholar .
Google Scholar .
Scelidosaurus harrisonii Owen, 1861 (Dinosauria: Ornithischia) from the Early Jurassic of Dorset, England: Biology and phylogenetic relationships.
Google Scholar .
Google Scholar B
Google Scholar B
The anatomy and palaeobiology of the early armoured dinosaur Scutellosaurus lawleri (Ornithischia: Thyreophora) from the Kayenta Formation (Lower Jurassic) of Arizona.
Google Scholar .
Scelidosaurus harrisonii Owen, 1861 (Dinosauria: Ornithischia) from the Early Jurassic of Dorset, England: Cranial anatomyB
Google Scholar .
Google Scholar .
(Ornithischia: Thyreophora) from the Lower Cretaceous Allaru Mudstone of Richmond, Queensland, Australia.
Google Scholar .
New ankylosaur (Thyreophora, ornithischia) remains from the Upper Cretaceous of Patagonia.
Google Scholar .
Google Scholar .
Google Scholar C
Dinosaur tracks in the Kokorkom Desert, Candeleros Formation (Cenomanian, Upper Cretaceous), Patagonia Argentina: Implications for deformation structures in dune fields.
Google Scholar .
High-resolution analysis of an erg-margin system from the Cretaceous Candeleros Formation (La Buitrera Paleontological Area, Río Negro Province, Argentina): An approach to different scales of fluvial-aeolian interactions.
Google Scholar .
Google Scholar .
Google Scholar .
Large Cretaceous sphenodontian from Patagonia provides insight into lepidosaur evolution in Gondwana.
Google Scholar .
New skulls and skeletons of the Cretaceous legged snake Najash, and the evolution of the modern snake body plan.
Google Scholar .
An early Late Cretaceous lizard from Patagonia, Argentina.
Google Scholar .
Google Scholar .
Highly specialized mammalian skulls from the Late Cretaceous of South America.
Google Scholar .
Google Scholar .
Google Scholar .
Google Scholar .
Google Scholar .
Google Scholar .
Google Scholar .
Google Scholar .
Google Scholar .
Basal Ceratopsia.
Google Scholar .
The skull of the basal stegosaur Huayangosaurus taibaii.
Google Scholar .
Google Scholar .
Google Scholar .
Google Scholar .
Scelidosaurus harrisonii Owen, 1861 (Dinosauria: Ornithischia) from the Early Jurassic of Dorset, England: Postcranial endoskeletonM
Google Scholar .
Euoplocephalus tutus and the diversity of ankylosaurid dinosaurs in the Late Cretaceous of Alberta, Canada, and Montana, USA.
Google Scholar .
Google Scholar J
Google Scholar .
Google Scholar .
Google Scholar B
A new ankylosaurid skeleton from the Upper Cretaceous Baruungoyot Formation of Mongolia: Its implications for ankylosaurid postcranial evolutionJ
Google Scholar .
A new basal ornithopod dinosaur (Frenchman Formation, Saskatchewan, Canada), and implications for late Maastrichtian ornithischian diversity in North America.
Google Scholar .
Scelidosaurus harrisonii Owen, 1861 (Dinosauria: Ornithischia) from the Early Jurassic of Dorset, England: Dermal skeleton.
Google Scholar .
Google Scholar .
Postcrania of juvenile Pinacosaurus grangeri (Ornithischia: Ankylosauria) from the upper Cretaceous Alagteeg Formation, Alag Teeg, Mongolia: Implications for ontogenetic allometry in ankylosaurs.
Google Scholar .
Google Scholar .
Google Scholar .
Google Scholar .
Google Scholar .
Google Scholar J
A reassessment of the purported ankylosaurian dinosaur Bienosaurus lufengensis from the Lower Lufeng of Yunnan, China.
Google Scholar .
Google Scholar C
Google Scholar .
Google Scholar .
Google Scholar .
Google Scholar .
New specimens of Scutellosaurus lawleri Colbert, 1981, from the Lower Jurassic Kayenta Formation in Arizona elucidate the early evolution of thyreophoran dinosaurs.
Google Scholar .
Google Scholar .
A new early branching armored dinosaur from the Lower Jurassic of southwestern China.
Google Scholar .
Google Scholar M
A basal thyreophoran (Dinosauria, Ornithischia) from the Tiouraren Formation of Niger.
Google Scholar .
Earliest record of an ankylosaurian dinosaur (Ornithischia: Thyreophora): dermal armor from the lower Kota Formation (Lower Jurassic) of India.
Google Scholar .
A new primitive neornithischian dinosaur from the Jurassic of Patagonia with gut contents.
Google Scholar .
New thyreophoran (Dinosauria, Ornithischia) remains from the lower Cretaceous Bajada Colorada Formation (Neuquén, Argentina).
Google Scholar .
Ornithischian remains from the Chorrillo Formation (Upper Cretaceous), southern Patagonia, Argentina, and their implications on ornithischian paleobiogeography in the Southern Hemisphere.
Google Scholar .
A new Cretaceous thyreophoran from Patagonia supports a South American lineage of armoured dinosaurs.


 UK
UK Australia
Australia France
France Germany
Germany Russia
Russia India
India Canada
Canada